Часть вторая
Развитие коацерватной жизни
ї 10 Развитие коацерватной жизни
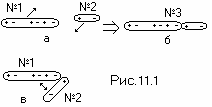
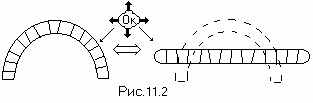
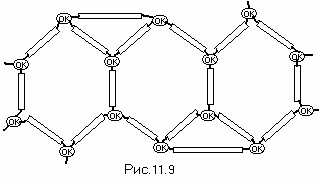
Замыкание ДНК в спираль
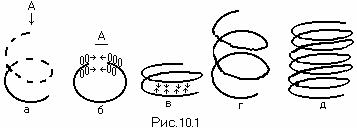
** **
** **
Обязательные свойства ДНК, как носителя информации и организация размножения
Атавизмы
Рудименты
** **
ї 11 Жизнь и окислители
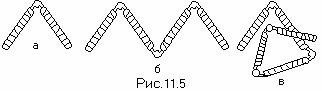
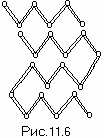
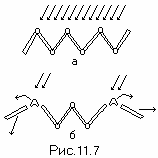
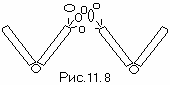
** **
** **
Об инстинктах
** **
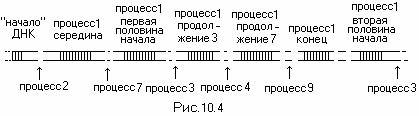
Скачки в записях в ДНК
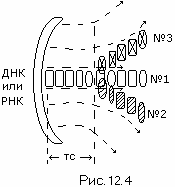
** **
Обязательные свойства жизни Таблица N1

Обязательные свойства жизни Таблица N1
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Для надежности и лучшего запоминания повторим таблицу в открытом виде.
- --
Замкнутое, клеточное строение.
- -- Обязательность периодического деления, размножения клетки на большой спиралевидной молекуле типа ДНК.
- -- Обязательность предохранения от преждевременного деления с использованием хромосомного чехла накопительных молекул типа РНК.
- -- Обязательность постоянных или периодических записей в ДНК, ее самопроизвольное удлинение, особенно в условиях длительного недоедания.
- -- Периодическое добавление дополнительных хромосом.
- -- Обязательное наличие клеточных молекулярных механизмов (КММ) для манипуляций с крупными молекулами.
- -- Обязательное наличие КММ для контроля ДНК на гладкость рабочей ее стороны и ремонта испорченных участков ДНК.
- -- Обязательность простановки меток отмены-переделки на ДНК.
- -- Обязательность периодического деления, размножения клетки на большой спиралевидной молекуле типа ДНК.
- --
Обязательное изменение организма по измененной наследственности в течение его собственной жизни. Наличие периодически действующих связей от организма в целом к ДНК и обратно.
- -- Инстинкты - программа готовых действий - записаны в организме в целом. В ДНК есть только косвенные указания того, как этот инстинкт реализуется в процессе роста организма.
- -- У белковой формы жизни - невероятное изобилие разных белков, синтезируемых без изменений в ДНК. Только скачкообразное изменение записей в ДНК в условиях длительного недоедания.
- -- Инстинкты - программа готовых действий - записаны в организме в целом. В ДНК есть только косвенные указания того, как этот инстинкт реализуется в процессе роста организма.